 |
 |
 |
 |
| Чемпионат УРФО по карпфишигу август 2013 г. |
 |
| Чемпионат России2011 Биг-Фиш |
 |
| Ловля сазана. Нижняя Волга |
 |
| Ловля сазана на реке Эбро |
 |
| Кубок Урала 2009 по карпфишингу |
 |
| Вести Сюжет про Кубок Урала по карпфишингу |
 |
| Кубок Регионов 2009 по карпфишингу |
 |
| Запуск карпа на озере Серебряное |
 |
| Фестиваль Уральских карпятников |
 |
| Кубок Урала 2008г. по карпфишингу |
 |
| Вести Карпфишинг по-южноуральски |
О восприятии цвета карпом
Что такое свет.
Перед тем, как начать эту статью, нужно определить некоторые, понятные нам с детства явления, воспринимаемые нами на уровне интуиции.
Основным таким понятием является свет. Свет это природное явление, одна из форм электромагнитного поля, воспринимаемая зрением. То есть свет это электромагнитное излучение, или иначе волна. Как всякая волна, свет излучается и поглощается физическими телами. Свет излучается нагретыми или иначе находящимися в возбужденном состоянии телами и веществами. Как физическое явление, Свет изучается в физике, в разделе оптика. Причем Свет это не одиночная волна с определенными характеристиками, а поток волн, разной длины и частоты. Из школьного курса физики мы знаем, что как всякие другие волны, свет может быть разложен на составляющие его волны при помощи дифракционной решетки (дифракция) или при помощи призмы (дисперсия). После такого разложения мы получаем спектр волн разной длины, при этом большой участок этого спектра будет невидим человеческим глазом. Дифракционный и дисперсионный спектры имеют некоторые различия. Дифракция, это явление отклонения от прямолинейного направления движения волны при прохождении ее через препятствия (щель, отверстие, стержень), размер которого соизмеримы с длиной волны. В случае дифракции, мы получаем картинку, имеющую несколько максимумов, не растянутую ни в какой из областей спектра (нормальный дифракционный спектр). Нормальный дифракционный спектр равномерный во всех областях и располагается в порядке возрастания длин волн. Дисперсия это физическое явление, связанное с распространением волн разной длины с разной скоростью в данном веществе. Коэффициент, полученный в результате таких опытов, называют коэффициентом преломления среды. Дисперсионный спектр сильно сжат в области волн имеющих большую длину, и сильно растянут в области волн имеющих меньшую длину волны. Дисперсионный спектр располагается в порядке убывания длин волн. Видимая часть спектра называется оптическим диапазоном спектра.
Цвет и спектральные цвета.
Что такое цвет? Физика дает следующий ответ на этот вопрос: Цвет, это качественная субъективная характеристика электромагнитного излучения оптического диапазона, определяемая на основании возникающего физиологического зрительного ощущения, и зависящая от ряда физических, физиологических и психологических факторов.
Индивидуальное восприятие цвета определяется его спектральным составом, а также цветовым и яркостным контрастом с окружающими источниками света и несветящимися объектами.
В непрерывном световом спектре, в котором одни цвета плавно переходят в другие так, что определить точно границы каждого цвета и связь его с определенной длиной волны сложно принято различать следующие цвета в зависимости от длины волны:
|
№ п/п |
Название цвета |
Длина волны (нм) |
||
|
От |
До |
|||
|
1 |
Фиолетовый |
380 |
440 |
|
|
2 |
Синий |
440 |
480 |
|
|
3 |
Голубой |
480 |
510 |
|
|
4 |
Зеленый |
510 |
550 |
|
|
5 |
Желто-зеленый |
550 |
575 |
|
|
6 |
Желтый |
575 |
585 |
|
|
7 |
Оранжевый |
585 |
620 |
|
|
8 |
Красный |
620 |
780 |
|
Диапазон волн от 0 нм до 380 нм, принято считать невидимым и называть ультрафиолетовой областью оптического излучения.
Диапазон волн от 780 нм до 1 мм, принято считать невидимым и называть инфракрасной областью оптического излучения.
Непрерывный оптический спектр
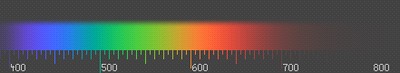
На рисунке 1 приведен главный максимум дифракционного цветового спектра.
Органы зрения живых существ воспринимают свет, отраженный от физических объектов и предметов. Цвет предмета, воспринимаемый органами зрения будет соответствовать длинам волн, отражаемых данными объектами. На пример, листва нам кажется зеленой по тому, что зеленую составляющую спектра лист отражает, а все другие составляющие, наоборот, поглощает. Или другой пример: апельсин оранжевый, по тому, что именно оранжевая составляющая светового спектра отражается апельсином.
Чувствительность органов зрения живых существ не постоянна в зоне видимого светового спектра. Для человека, на пример, на основании данных [3.2] чувствительность органов зрения приведена на Рисунке 2.
Спектральная чувствительность палочкового зрения (рисунок 2, кривая 2 – глаз адаптирован к ночным яркостям) характеризует работу глаза при столь малом количестве света, что его не хватает даже для частичного возбуждения колбочек. Кривая относительной спектральной чувствительности глаза имеет максимум на длине волны в 507 нм.
Для глаза, адаптированного к дневным яркостям V(λ) (рисунок 2, кривая 1), на длинах волн 510 нм и 610 нм характерно двукратное снижение чувствительности. Если же глаз адаптирован к ночным яркостям V’(λ) (рисунок 2, кривая 2), то снижение чувствительности в два раза наблюдается на длинах волн 455 нм и 550 нм.
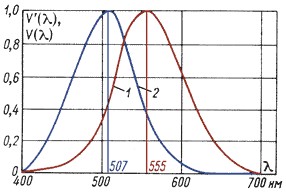
Рисунок 2. Относительная спектральная чувствительность глаза человека
Максимумы на кривых 1 и 2 на рисунке 2, равные единице, относительны. Дело в том, что палочковый аппарат ночного зрения человека намного чувствительнее, и для восприятия предельно малого светового сигнала (например, едва видимой точки на темном фоне) палочкам необходима примерно в пятьсот раз меньшая мощность, чем колбочкам. При этом палочки, действующие при периферическом (боковом) зрении, не позволяют определить цвета точки, в то время как колбочки, фиксирующие точку при прямом зрении, дают возможность увидеть и ее цвет.
Кроме этого, чувствительность человеческого глаза неодинакова к разным цветовым компонентам света. Чувствительность максимальна при 555 нм (желто-зеленый свет) и сводится к минимуму при более длинных (красный свет) и коротких (синий свет) длинах волн. Чувствительность человеческого глаза к воздействию красного излучения (650 нм) составляет всего 10% от максимальной чувствительности. Иными словами, чтобы добиться ощущения той же яркости, что и у желто-зеленого света, интенсивность красного света должна быть в десять раз больше.
Если соединить видимые красный и синий диапазон спектра, то мы получим цветовой круг Рисунок 3. Цветовой круг это способ представления непрерывности цветовых переходов в видимой части спектра. Сектора круга окрашены в различные цветовые тона, размещенные в порядке расположения спектральных цветов, причем пурпурный цвет связывает крайние красный и фиолетовый цвета.

Рисунок 3. Цветовой круг и триады цветов, дающие при смешивании белый цвет.
Цветовой круг впервые был предложен Исааком Ньютоном в 1704 году. Цветовой круг имеет большое значение для понимания законов смешивания спектральных цветов. Так на пример, вершины треугольника, вписанного в цветовой круг, однозначно указывают на триады цветов, которые при смешивании дадут белый цвет.

Рисунок 4. Цветовое поле видимого спектра.
В общем случае, оттенки цветов получаемые при смешивании простых спектральных цветов представлены на Рисунке 4.
Не спектральные цвета и смешивание цвета.
Для восприятия цвета очень важно такое явление, как метамерия, особенности глаза и психики. Метамерия, это свойство зрения, при котором свет различного спектрального состава может вызывать ощущение одинакового цвета. Иначе метамерией можно назвать восприятие двух окрашенных образцов одинаково окрашенными под одним источником освещения, но различно окрашенными под другим источником освещения. Это можно объяснить разными спектральными характеристиками источников освещения и разными наполняющими цветами в красочных покрытиях рассматриваемых образцов.
Физиологически метамерия зрения основана на строении периферического отдела зрительного анализатора биологического объекта. В соответствии с теорией происхождения видов, предки человека получили органы зрения от рыб. Эта гипотеза получила в настоящее время, как множество подтверждений, так и не меньшее число опровержений.
У человека, как и у карпа, роль периферического отдела зрительного анализатора выполняет сетчатка, в которой за восприятие цвета отвечают особые клетки, называемые колбочками.
В общем случае, можно создать такие условия, при которых пучок оранжевого спектрального цвета, пучок оранжевого не спектрального цвета (полученный смешением желтого и красного спектральных цветов) и пучок пурпурного не спектрального цвета (полученный смешением синего и красного спектральных цветов) могут восприниматься зрительным анализатором наблюдателя, как пучки одинакового цвета.
Однако если пропустить эти три пучка через дисперсионную призму, то мы получим:
Для оранжевого спектрального цвета: одну полоску, соответствующую длине волны первичного светового пучка.
Для оранжевого не спектрального цвета (полученного смешением желтого и красного спектральных цветов): две полоски, соответствующие длинам волн составляющих желтого и красного спектральных цветов первичного светового пучка.
Для пурпурного не спектрального цвета (полученного смешением синего и красного спектральных цветов): две полоски, соответствующие длинам волн составляющих синего и красного спектральных цветов первичного светового пучка.
В общем случае, результирующие цвета получаемые при смешивании цветов иллюстрирует Рисунок 5.

Рисунок 5. Результирующие цвета, получаемые при смешивании спектральных цветов
Данное наблюдение представляется мне важным при создании цвета красителя для окрашивания насадки.
Теории восприятия цвета.
На сегодняшний день, существуют несколько теорий восприятия цвета. Пожалуй, самой распространенной из них является Трехкомпонентная теория, предложенная тремя авторами: М.В. Ломоносовым, Т. Юнгом и Г. Гельмгольцем. Согласно этой теории, в органе зрения человека существуют три цветоощущающих аппарата: красный, зеленый и синий. Каждый из них возбуждается в большей или меньшей степени, в зависимости от длины волны излучения. Затем возбуждения суммируются аналогично тому, как это происходит при суммируемом смешении цветов. Суммарное возбуждение ощущается человеком как тот или иной цвет. В своей работе «Цветовое зрение» авторы Л.Н. Миронова, И.Д. Григорьевич отмечают: «…Трехкомпонентная теория хорошо объясняет важнейшие закономерности цветового зрения: адаптацию, индукцию, цветовую слепоту, спектральную чувствительность глаза, зависимость цвета от яркости и другие, Однако, следует заметить, что в наше время известны факты, свидетельствующие о более сложной картине функционирования органа зрения…».
Другой, очень распространенной и имеющей множество подтверждений, теорией является теория оппонентных цветов Э. Геринга. Геринг выдвинул предположение, что в колбочках сетчатки могут существовать три вида гипотетических веществ: бело-черные, красно-зеленые и желто-синие. Световой поток влечет их разрушение (одни световые лучи) с образованием белого, красного или желтого цветов или синтез (другие световые лучи) чорного, зеленого или синего цвета. Геринг предполагал, что имеются четыре основных цвета красный, желтый, зеленый и синий, и что они попарно связаны с помощью двух антагонистических механизмов зелено-красного механизма и желто-синего механизма. Постулировался также третий оппонентный механизм для ахроматически дополнительных цветов белого и черного. Из-за полярного характера восприятия этих цветов Геринг назвал эти цветовые пары «оппонентными цветами». Из его теории следует, что не может быть таких цветов, как зеленовато-красный и синевато-желтый. Таким образом, теория оппонентных цветов постулирует наличие антагонистических цветоспецифических нейронных механизмов. Например, если такой нейрон возбуждается под действием зеленого светового стимула, то красный стимул должен вызывать его торможение. Предложенные Герингом оппонентные механизмы получили частичное подтверждение после того, как научились регистрировать активность нервных клеток, непосредственно связанных с рецепторами. Так, у некоторых позвоночных, обладающих цветовым зрением, были обнаружены красно-зеленые и желто-синие горизонтальные клетки. У клеток красно-зеленого канала мембранный потенциал покоя изменяется и клетка гиперполяризуется, если на ее рецептивное поле падает свет спектра 400-600 нм, и деполяризуется при подаче стимула с длиной волны больше 600 нм. Клетки желто-синего канала гиперполяризуются при действии света с длиной волны меньше 530 нм и деполяризуются в интервале 530-620 нм.
Множество проводимых исследований подтвердили предположения этих двух теорий, так например колбочки у приматов существуют всего трёх типов: воспринимающие цвет в фиолетово-синей, зелено-жёлтой, в желто-красной частях спектра. Каждый вид колбочек интегрирует поступающую лучистую энергию в довольно широком диапазоне длин волн, и диапазоны чувствительности трёх видов колбочек перекрываются, различаясь лишь диаграммой величины чувствительности.
Человеческое зрение, таким образом, является трёхстимульным анализатором, то есть спектральные характеристики цвета выражаются всего в трех значениях. Если сравниваемые потоки излучения с разным спектральным составом производят на колбочки одинаковое действие, цвета воспринимаются как одинаковые.
В животном мире известны четырёх- и даже пятистимульные цветовые анализаторы, так что цвета, воспринимаемые человеком одинаковыми, животным могут казаться разными так, хищные птицы видят следы грызунов на тропинках к норам исключительно благодаря ультрафиолетовой люминисценции компонентов их мочи.
Характеристика органов зрения карпа.
Как уже говорилось выше, в соответствии с теорией происхождения видов, предок человека унаследовал органы зрения от низших позвоночных, или от рыб, что вызывает большое сомнение у некоторых, уважаемых в научных кругах, авторов:
«…Если принять как факт, что цветовое зрение мы унаследовали от низших позвоночных (рыб), что доказывается анатомическим, физиологическим, химическим и структурным сходством строения сетчатки, то эволюцию цветового зрения следует изучать не на приматах, а начиная с рыб. Тогда рассуждения об эволюции цветового зрения от протонопии до тритонопии (С.В.Кравков) нельзя признать обоснованной. Ведь уже у карпа имеются все три типа колбочек и даже детекторов оппонентного типа, хотя и находятся эти детекторные клетки еще в самой сетчатке, а не в латеральном коленчатом теле, как у приматов и человека (Пэдхем Ч., Сондерс Ж., 1978). Хотя карп обладает повышенной чувствительностью в красно-оранжевой, а не зелено-желтой области спектра, диапазоны частот реагирования рецепторов карпа и человека почти не различаются по ширине.
Таким образом, эволюция цветового анализатора шла параллельно с развитием анатомических и функциональных отделов центральной нервной системы (промежуточного, среднего мозга, коры), по «вертикали», а не в сторону дифференциации цветочувствительных клеток периферического отдела анализатора (колбочек сетчатки), по горизонтали. Сохранение (даже небольшое расширение) диапазона чувствительности при усовершенствовании структуры цветового анализатора в ходе наземной эволюции высших позвоночных свидетельствует, что цвет играл существенную роль в их жизнедеятельности. Но только у той биологической линии, которая привела к возникновению человека. О центральном значении цветоразличения для человека свидетельствует хотя бы тот факт, что все 6,5 миллиона колбочек как у карпа, так и у человека, располагаются в фовеа-центральной зрительной ямке, области максимально четкого зрения …».
Далее автор делает вывод: «…Итак, мы видели, что для видов, значительно уступающих человеку в психическом развитии (растения, насекомые, рыбы, пресмыкающиеся, птицы), цвет не отделим от функций размножения, питания и выживания, т.е. от всего биологического цикла …».
Из сказанного становится ясным, что максимум цветового восприятия карпа обыкновенного лежит в красно-оранжевой области светового спектра, что находит свое подтверждение в большом количестве работ других авторов.
Автор считает, что диапазон чувствительности в области видимого спектра карпа обыкновенного и человека почти не различается по ширине, что противоречит данным некоторых других источников.
Автор подчеркивает подобность строения органов зрения карпа и человека не только качественно: «…колбочки как у карпа, так и у человека располагаются в фовеа-центральной зрительной ямке…» но и количественно: «…все 6,5 миллиона колбочек как у карпа, так и у человека…».
Кроме этого, автор считает функцию распознания цвета, в частности карпом, не отделимой от функций размножения, питания и выживания, то есть от всего биологического цикла. Это пожалуй самый ценный для нас вывод, для нас – рыбаков, осуществляющих ловлю этой умной рыбы, маскируя насадку под привлекающую ее, рыбу, пищу.
Справедливости ради, нужно отметить, что в результате биофизических исследований органов зрения рыб, пресноводных, пресмыкающихся, приматов, человека, были получены другие, очень интересные факты, способные поставить под сомнение приведенные выше заключения.
Так на пример, в своей статье «О зрении животных», опубликованной в электронном журнале «LiveJournal» некий Евгений [6], приводит интересные факты, касающиеся возможностей органов зрения различных животных, птиц, рыб, ссылаясь на результаты научных исследований, опубликованные в научных изданиях. Так на пример, относительно золотых рыбок, относящихся к карповидным, Евгений пишет: «… 14. Золотые рыбки – тетрахроматы и видят длины волн от 300 нм (и даже ниже) до примерно 730 нм – то есть весь человеческий диапазон, плюс хороший кусок ультрафиолета, плюс пограничную с инфракрасным область… ». Ссылаясь на статью известных биологов Shozo Yohoyama, Huan Zhang, Z. Bernhard Radlwimmer, Nathan S. Blow «Adaptive Evolution of Color Vision of the Commoran Coclacanth (Latimeria Chalumnae)», опубликованную в 26 марта 1999 года в Ню-Йорке, и перепечатанную журналом «Evolution» в мае 1999 года, Евгений пишет: «…15. Латимерия (целакант) – древняя, долгое время считавшаяся вымершей рыба, обитающая на глубине около 200 м. Света там почти нет, а те его остатки, что всё-таки туда просачиваются – исключительно синие. Тем не менее, она тоже обладает цветным зрением, с нашей точки зрения весьма уникальным. Латимерия – дихромат, но всё богатство воспринимаемой ею гаммы укладывается, по нашим меркам, в почти неотличимые оттенки синего в узеньком диапазоне длин волн возле 480 нм. Максимумы цветового восприятия её рецепторов отстоят друг от друга всего на 7 нанометров: 478 и 485 нм. …». Относительно карпа обыкновенного, ссылаясь на статью «The eyes of the common carp and Nile tilapia are sensitive to near-infrared» японских авторов Taro MATSUMOTO and Gunzo KAWAMURA, Евгений пишет: «…16. Обыкновенный карп может видеть в ближнем инфракрасном диапазоне (865 нм) – там же, где работают пульты управления телевизором и где рассеяние света в воде и воздухе существенно ниже. …».
Учитывая эти данные, мы можем усомниться в утверждении о равенстве ширины светового спектра воспринимаемого органами зрения карпа и человека.
Интересным представляется мнение А.М. Черноризова, высказанное им в его докторской диссертации на тему «Нейронные механизмы цветового зрения».
Проведя опыты на речном карпе, Carpio Cyprims L.; на 13 карпах in vivo (живых карпах) и более чем 200 карпах in vitro, и проанализировав другие, известные ему работы, автор пришел к заключению: «…Исследование и моделирование процессов передачи информации о цвете в нейронных сетях зрительной системы является одной из главных задач психофизиологии цветовосприятия в рамках современной психофизиологии как науки о нейронных механизмах психических процессов и состояний. Адекватной экспериментальной моделью для этого является сетчатка глаза, которая по сложности строения и возможностям интегральной обработки параметров зрительных образов напоминает мозг (Jasper, Raynauld, 1975; Хьюбел, 1990). На уровне нервных элементов сетчатки осуществляется переход от кодирования цвета цо принципам трехкомпонентной теории Ломоносова-Юнга-Гельмгольца (слой фоторецепторов) к кодированию цвета по принципам теории оппонентных цветов Геринга (слой горизонтальных и биполярных клеток). В сетчатке происходит формирование цветооппонентных («красно-зеленых», RG-типа; "сине-желтых", YB-типа) и ахроматических (нецветооппонентных «яркостных», В-типа, и "^темновых", D-типа) нейронных систем, которые, по нашим данным, обладают разной функцией в процессе цветокодирования. …».
Далее автор отмечает: «…В поведенческих опытах Wolf (1925) показано, что рыбы из одного с карпом семейства Cyprinidae могут различать до 20 различных цветов в диапазоне от 340 до 760 нм. При этом рыбы отличают пурпурный цвет (смесь синего и красного спектральных излучений) от любого другого цвета. Hamburger (1926) выявил существование дополнительных цветов для рыб {Phoxinus laevisAG, семейство Cyprinidae), а также способность отличать белый цвет от какого-либо спектрального цвета. Таким образом, всю гамму цветов для рыб, как и для человека, можно представить в виде замкнутой круговой диаграммы (круга Ньютона) (Herter, 1953). Herter (1953) констатировал явления одновременного и последовательного яркостного и цветового контрастов для цветового зрения рыб. Horio (1938) в опытах на карпах показал, что при различении зрительных стимулов рыбы чаще ориентируются на цвет, чем на форму. Способность рыб, в частности, карпа, правильно оценивать цвет предметов независимо от условий освещения (константность восприятия цвета) продемонстрирована в поведенческих и Электрофизиологических исследованиях (Oyama, Jitsumori, 1974; Диментман и др., 1975; Максимова и др., 1975; Crawford et.al., 1990). Наконец, цветовое зрение рыб, как и у человека, трихроматично. На это указывают данные микроспектрофотометрических, нейрофизиологических и поведенческих экспериментов (для обзора см.: Измайлов и др., 1989).
Имеются данные о наличии в сетчатке костистых рыб фоторецепторов с пиком чувствительности в ультрафиолетовой области спектра (Neumeyer, Arnold, 1989). Не ясна роль этих рецепторов в цветовом зрении рыб ввиду того, что оптическая система камерного глаза этих животных не пропускает ультрафиолетовые лучи. Однако, имеются данные о влиянии активности рецепторов этого типа в различение цветов в синей области спектра (400-480 нм) (Neumeyer, Arnold, 1989).
У рыб и амфибий хорошо развита система ретино-тектальных зрительных проекций, что обусловливает сложный характер обработки цветового сигнала уже на уровне нейрональных структур сетчатки. В этой ситуации сетчатка этих животных может служить моделью для изучения принципов цветокодирования, реализуемых у приматов центральными отделами зрительного анализатора. …»
Подводя итог проведенным исследованиям, автор замечает: «…Достоверность результатов достигалась большим объемом выборки и использованием современных статистических методов многомерного анализа (метрическое многомерное шкалирование). Представленные в работе данные получены в более чем 500 опытах на 26 моллюсках, 40 лягушках, 13 карпах in vivo и более чем 200 карпах in vitro. На изолированной сетчатке карпа внутриклеточно исследованы спектральные реакции 538 горизонтальных клеток и 45 биполярных клеток. …».
Анализируя приведенные выше исследования, с высокой степенью достоверности, можем предположить следующее:
Сетчатки глаза карпа и человека очень похожи по функционированию и строению, и «…по сложности строения и возможностям интегральной обработки параметров зрительных образов напоминают мозг…».
Имеются данные о наличии у рыб рецепторов, помогающих им различать цвета в области синего цвета, а также в значительном диапазоне инфракрасной зоны спектра.
Особое внимание нужно обратить на то, что максимальная активность нейронов мозга карпа зарегистрирована при раздражении его фоторецепторов пурпурным цветом, который является не спектральным цветом, а результатом суммирующего действия двух спектральных цветов: синего и красного.
Выводы.
1. Сетчатки глаза карпа и человека очень похожи по функционированию и строению, а по сложности строения и возможностям интегральной обработки параметров зрительных образов напоминают мозг.
2. Диапазон чувствительности в области видимого спектра органов зрения карпа и человека значительно различается по ширине, что дает карпу возможность видеть объекты в синей части спектра и в невидимой области инфракрасного цвета, предположительно до длин волн около 865 нм. Это, в свою очередь, объясняет то, как карп может найти пищу в условиях практически полной темноты, например, ночью.
3. Максимум цветового восприятия карпа лежит в красно-оранжевой области светового спектра.
4. Максимальная активность нейронов мозга карпа зарегистрирована при раздражении его фоторецепторов пурпурным цветом, который является не спектральным цветом, а результатом суммирующего действия двух спектральных цветов: синего и красного.
5. Карп способен отличать белый спектральный цвет от какого-либо другого цвета.
6. Во время распознавания объекта, карп более склонен ориентироваться на цвет объекта, чем на его форму.
7. Функцию распознания цвета у карпа не отделима от функций размножения, питания и выживания, то есть от всего биологического цикла.
Заключение.
Сделанные мною и приведенные выше, выводы, не претендуют на научную ценность и вполне могут быть ошибочными. Но в своих экспериментах с окрашиванием насадок я придерживаюсь следующих, изложенных мною ниже правил.
Мои насадки имеют преимущественно оранжевый спектральный цвет, оранжевый не спектральный цвет, пурпурный не спектральный цвет и белый не спектральный цвет. Они обязательно яркие и отчетливо выделяются на фоне окружающих предметов.
Насадки дают обильное, легко различимое облако мути имеющее тот же цвет, что и насадка.
В свои насадки я пытаюсь включать энзимы, способные поднять температуру поверхности насадки, по сравнению с температурой окружающей среды, хотя бы на один градус. Это позволяет выделить насадку на фоне окружающих ее объектов (заставляет насадку светиться изнутри) и делает ее более привлекательной и легко распознаваемой рецепторами инфракрасного зрения зрительного аппарата рыбы.
Автор Сиваченко Григорий.
Источник: www.sportfishing.ua